




 Найти эксперта
Найти эксперта

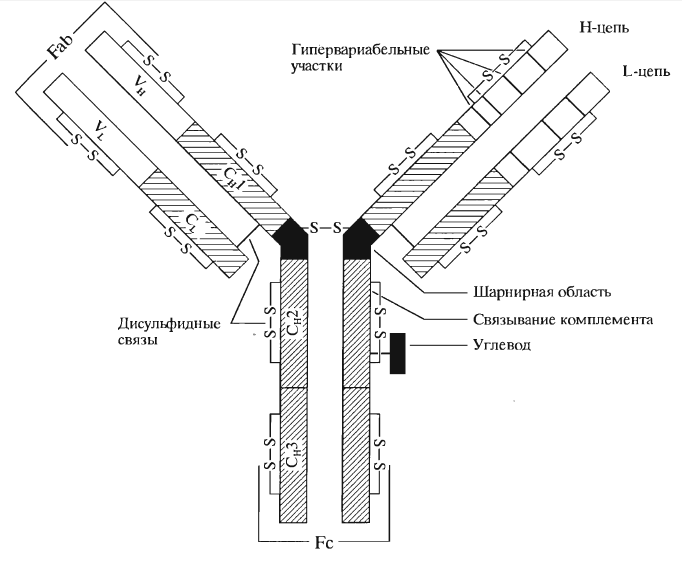
Определение. Структура иммуноглобулинов
Иммуноглобулины ($Ig$) – это класс структурно связанных белков гликопротеинв, состоящих из двух видов парных полипептидных цепей:
- легкие цепи ($L$), имеющие низкую молекулярную массу (25 кДа);
- тяжелые цепи ($H$), с высокой молекулярной массой (50-70 кДа).
Все цепи между собой дисульфидными мостиками.
Существует два типа легких цепей:
- каппа,
- лямбда.
Фермент папаин гидролизует молекулу с образованием двух идентичных антиген-связывающих $Fab$-фрагментов и одного не связывающего антиген $Fc$-фрагмента, или кристаллизующегося фрагмента. На этом фрагменте находятся участки связывания фрагмента комплемента $C1g.$
Легкие цепи имеют две протяженные области примерно равного размера:
- константная область во всех иммуноглобулинах приблизительно одинакова;
- вариабельная область, в ней аминокислотная последовательность очень изменчива.
В состав константной и вариабельной областей входит около 110 аминокислот.
Тяжелые цепи состоят из одной вариабельной области и трех или четырех ($IgM$, $IgE$) константных областей.
Для отдельных доменов молекулы иммуноглобулина характерна сходная глобулярная структура, сформированная множеством $β$-слоев и дисульфидных мостиков.
В состав вариабельных областей легких и тяжелых цепей входят участки с наиболее изменчивой аминокислотной последовательностью – гипервариабельные области.
Гипервариабельные области определяют специфичность связывания антител и являются областями, определяющими комплементарность к антигену. Замена одной аминокислоты в этой области может критически сказаться на связывании определенного антигена.
Константная область определяет эффекторную функцию иммуноглобулина – взаимодействие с Fc-рецепторами (специфическими рецепторами) различных клеток, степень связывания комплемента, перенос через плаценту.
Схема строения иммуноглобулина
Иммуноглобулины способны экспрессироваться на поверхности зрелых $B$-лимфоцитов и служат рецепторами антигенов. Отдельные эндогенные иммуноглобулины могут связываться с другими клетками (тучными клетками, гранулоцитами, эпителиальными клетками, моноцитами/макрофагами) с помощью $Fc$-рецепторов.
Окончательно дифференцированные $B$-клетки синтезируют рецепторы, которые секретируются в кровь как антитела.
Классы иммуноглобулинов
Все иммуноглобулины на основании антигенных и структурных признаков тяжелых пептидных цепей, в порядке относительного содержания в сыворотке крови, делят на пять классов:
- $IgG$ – 80%;
- $IgA$ – 15%;
- $IgM$ – 10%;
- $IgD$ – менее 0,1%;
- $IgE$ – менее 0,01%.
Молекулы $IgG$, $IgD$, $IgE$ являются мономерными, $IgM$ – пентамер, молекулы $IgA$ могут быть мономерами (сыворотка крови) или димерами (экскретируемые жидкости: слюна, слезная жидкость, секреты слизистых оболочек).
Иммуноглобулины могут выступать в роли антигенов. Они имеют следующие детерминанты:
- изотипические;
- аллотипические;
- идиотипические.
У иммуноглобулинов классов $IgG$ и $IgA$ выделяют несколько подклассов.
Классы и подклассы называют изотипами, которые одинаковы у всех особей одного вида.
Аллотипы – это индивидуальные аллельные варианты в пределах одного изотипа.
Антитела по антигенной специфичности относятся к различным идиотипам. Идиотипические детерминанты отвечают за вариабельность $CDR$-области определенной молекулы иммуноглобулина.
Циркулирующие антитела образуются и секретируются плазматическими клетками в:
- костном мозге,
- лимфатических узлах,
- ассоциированной со слизистой лимфоидной ткани.
Иммуноглобулин $IgA$ преобладает в:
- бронхиальных секретах,
- слюне,
- слезной жидкости,
- секретах мочевыводящих путей,
- грудном молоке и молозиве.
Переключение класса иммуноглобулинов
В процессе иммунного ответа синтезируются иммуноглобулины разных классов:
- зрелые В-клетки синтезируют $IgM$;
- перегруппировавшиеся $VDJ$-последовательности соединяются с другими $C$-генами константной области;
- переключающаяся $S$-последовательность контролирует процесс перегруппировки путем рекомбинации с другими $S$-последовательностями, которые обладают высоким уровнем гомологии;
- происходит делеция последовательностей генов константной области, расположенных между $VDJ$-последовательностями и новыми $C$-генами.
Найти эксперта



